



SEXUAL REPRODUCTION IN FLOWERING PLANTS
INTRODUCTION
- The process of reproduction takes place in the plant through a special structure, called flower.
- Calyx of sepals, corolla of petals, androecium of stamens and gynoecium of carpels are present in the typical or complete flower.
- The calyx and corolla are termed accessory whorls of the flower, because these structures do not participate in the process of reproduction, but only helps.
- The androecium and gynoecium are known as essential whorls, because they are directly related with the reproduction.
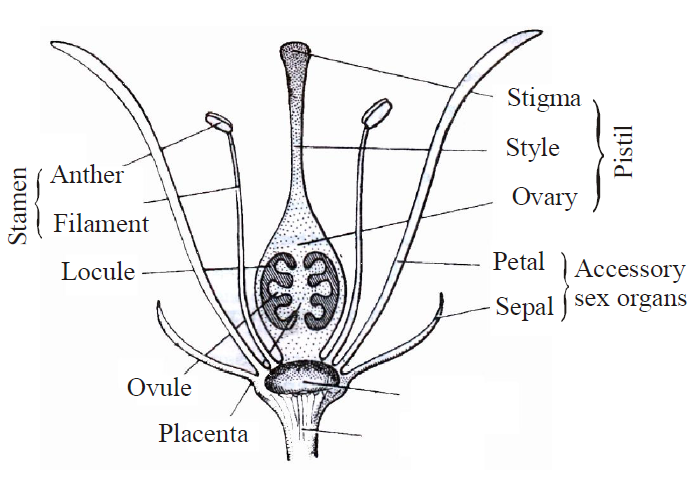
Fig. : Structure of Flower
- First of all N.Grew realized the fact, stamens are male sex organ of flower (anatomy of plants).
- Sexuality in plant was first reported by Jacob Camerarius.
- He reported anthers are the male sex organ and ovary with style and stigma are female sex organ, and for the formation of seed, interaction is essential in between both the sex organs.
- Significance of pollination and role of insects in pollination was recognized by Josheph Kolreuter.
- C.F. Wolf is the Father of plant embryology.
Prof. P. Maheshwari is Father of Indian plant Embryology.
He wrote a book - 'An Introduction to Embryology of Angiosperms'.
SEXUAL REPRODUCTION
- Sexual reproduction is the process of development of new organisms through the formation and fusion of gametes. In flowering plants, stamens are male reproductive organs while carpels are female reproductive organs.
- In angiosperms, male and female gametes are formed in male and female sex organs by the process of meiosis. Both the gametes fuse together to form a diploid zygote which gives rise to embryo.
- The process by which the embryo is formed by meiosis and fertilization is called amphimixis.
- The study of sexual reproduction in flowering plants includes pre-fertilization structures & events and post-fertilization structures & events.
- Pre-fertilization events includes male reproductive organ (androecium), female reproductive organ (gynoecium), pollination and fertilization.
Post-fertilization events includes development of endosperms, development of embryo (in dicot and monocot) and seed.
MALE REPRODUCTIVE ORGAN - ANDROECIUM
- Male reproductive organ is called androecium and their unit is called stamen.
Stamen is also known as microsporophyll. There are 6 stamens in Capsella. A typical stamen differentiates into three parts -a long, thin structure, called filament which joins the stamen to the thalamus. The free end of the filament, a swollen spore bearing structure is called anther. Anther and filament are attached together with the help of small region, called connective. Connective contains vascular tissues.
- Each anther is generally a bilobed structure i.e., anther has two anther lobes and is called dithecous.
- Each lobe of anther has two chambers which are called pollen sacs or pollen chambers.
Therefore, a typical anther has four pollen sacs (called tetrasporangiate).
- Pollen grains are formed inside the pollen sac through the meiotic division of pollen mother cells.
- At the maturity of the pollen grains, sterile tissue degenerate which are present in between the pollen sacs. Both the pollen sac are fused together. Because of this reason, only one chamber appears in each anther lobe at maturity. So two chambers are seen in the mature anther at the time of dehiscence.
- In Malvaceae, the anther of stamen has only one anther lobe. This is called monothecous and it contains only two pollen sacs called bisporangiate.
Monothecous anther are also found in Moringa, Wolffia plants.
When there is only one microsporangium per anther (e.g., Arceuthobium) then this condition is called monosporangiate.
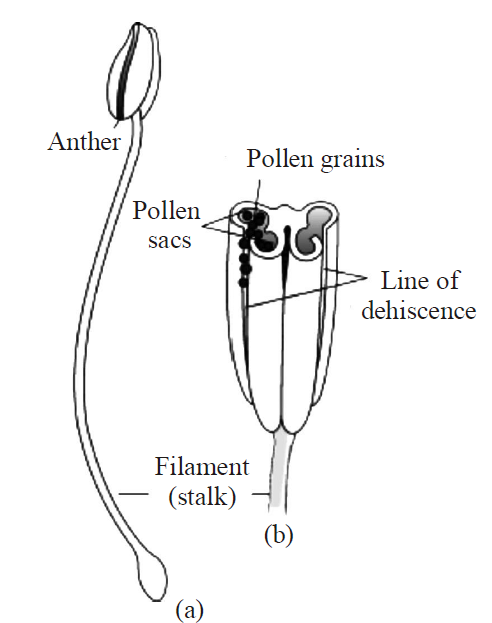
Fig. : (a) A typical stamen;
(b) three–dimensional cut section of an anther
STRUCTURE OF ANTHER
- The development of anther in origin is Eusporangiate type i.e. it is developed from more then one archesporial cells.
- In the transverse section of anther, it is seen almost spherical.
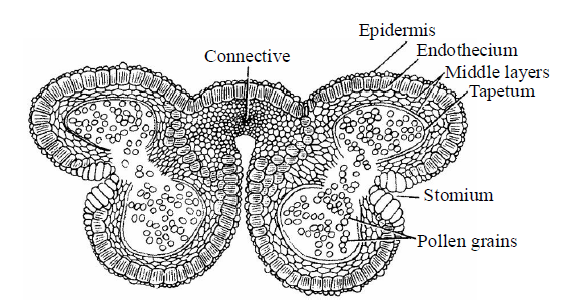
Fig. : T.S. of mature dithecous anther
- Anther consists of the following wall layers – epidermis, endothecium, middle layer and tapetum.
EPIDERMIS
- It is the outermost protective layer of anther. It is single celled thick and continuous layer but not archesporia in origin.
- In Arceuthobium (smallest parasitic angiosperm), fibrous thickening is present in epidermis so it is called exothecium.
ENDOTHECIUM
- This layer is present below the epidermis. It is single celled thick layer. During the maturation of anther, various changes take place in different walls of cells of endothecium. The outer wall of these cells remains thin walled, but inner walls and radial walls become thick due to thickening of α-cellulose fibers.
- Callose bands are also present along the radial walls. At some places, callose bands and fibrous thickening are absent. These places are called stomium. The dehiscence of anther takes place only from these places. Endothecium becomes hygroscopic due to the presence of fibrous thickening. So, it helps in dehiscence of anther.
NOTES
In Hydrocharitaceae family, fibrous thickening is absent in endothecium.
MIDDLE LAYER
- Middle layer consists of parenchymatous cells. This layer is one to three celled thick structure. Food is stored by parenchymatous cells in this layer.
- Middle layer is ephemeral in nature and absent in a mature anther.
NOTES
- In Holoptelea plant, 3 to 4 celled thick middle layer is present.
- In Najadaceae & Lemnaceae families, middle layer is absent.
- In Wolffia, middle layer is absent.
TAPETUM
- It is the single celled inner most thick layer which acts as nutritive layer.
- Pollen sacs are surrounded by tapetum.
- The cells of the tapetum are initially diploid but they become polyploid due to endomitosis. It means these cells contain many chromosomes.
- Tapetum absorbs food from the middle layer and provide nutrition to the microspore mother cells or microspores. The cells of tapetum secrete hormones and callase enzymes. The tapetum layer disappears in the mature anther.
NOTES
In Nicodia and Costum plants, tapetum is multilayered.
- The tapetal cells play a significant role during the meiotic cell division in micro sporogenous cells and in pollen development.
- Tapetum is of two types - amoeboid and secretory.
Amoeboid Tapetum/Invasive tapetum/Periplasmodial tapetum: It is found in primitive angiosperms. Such type of tapetum absorb all foods from the middle layer. So middle layer immediately degenerates. In the beginning, all food materials are stored by tapetum. Tapetal cells convert absorbed food into special food granules called protoplast bodies. The innermost layer of tapetum dissolve and release its protoplast into the cavity of the microsporangium. Now inside the pollen sacs protoplast bodies are known as periplasmodium. Microspore mother cells are surrounded by
periplasmodium and provides nourishment to the developing microspores. This type of tapetum provide nutrition to the microspores after degeneration .
periplasmodium and provides nourishment to the developing microspores. This type of tapetum provide nutrition to the microspores after degeneration .
Example: Typha, Alisma and Tradescantia.
Glandular or Secretory Tapetum : It does not degenerates quickly. Cells of this tapetum remain intact throughout the microspore development. It absorbs nutrients from the middle layer and secrete into the cavity of the microsporangia (pollen sacs) and does not store it. E.g., most of flowering plants (Capsella).
Before degeneration of cells of tapetum, they form special granules called Pro Ubisch bodies in the cytoplasm. Pro Ubisch bodies transfer between cell wall and cell membrane of tapetal cells. Here, they are surrounded by sporopollenin. Now they are called Ubisch bodies or orbicules. At last, tapetum degenerates and ubisch bodies are released into pollen sacs. Ubisch bodies participate in the formation of exine of the microspores inside the pollen sacs. Generally, sporopollenin participates in the formation of outer covering (exine) of pollen grains.
NOTES
Tapetum helps in transfer of food, storage of food, formation of sporopollenin and pollen kit materials.
MICROSPOROGENESIS
- The formation of microspores or pollen is called microsporogenesis : Pollen grains are formed inside the anther.
- The anther appears as outgrowth like structure in the initial stage which shows spherical or oval shaped structure.
- At this stage, it is a mass of undifferentiated and homologous meristematic cells which is surrounded by a single cells thick outer layer called the epidermis. First of all vascular tissue are formed in middle region. Simultaneously, four cells located just below the epidermis in vertical rows in the region of hypodermis at the four corners become large. It has visible nucleus with dense cytoplasm. Due to this reason, they are different from the rest of the cells. These cells are called archesporial cells.
- Archesporial cells divide periclinal to form primary parietal cells below the epidermis and primary sporogenous cells towards the centre. Both of the cells usually undergo further divisions to form complete structure of anther except epidermis.
- Primary parietal cells undergo further periclinal and anticlinal division to form a series of 3-5 layers making the walls of the anther.
- The primary sporogenous cells divide twice or more than two by mitotic division to form sporogenous cells and later sporogenous differentiates into microspore mother cells (MMC) during the formation of the wall of the pollen sac.
- Each MMC divide to form four haploid microspore or pollen grain by meiotic division or reduction division.
- At the initial stage, all four microspores are attached together with the help of callose layer. This group of microspores is called tetrad. After some time, this callose layer dissolve by callase enzyme, which is secreted by tapetum.
- Normally each microspore mother cell can form tetrad by meiotic division. But in some plant like Zostera, some microspore mother cells become sterile and provide nutrition to rest of microspore mother cells.
- Similarly, tapetum is not well developed in Gentianaceae family so some cells of sporogenous tissue become sterile and provide nutrition to remaining sporogenous cells.
Types of tetrads :The arrangement of the microspores in tetrad condition are as follows:
- Tetrahedral tetrad (most common) : Four haploid microspores arranged in tetrahedral form.
E.g., Dicotyledons- (Capsella)
- Isobilateral tetrad : This condition is found in monocotyledons. Microspores are arranged at the lateral side of each other.
- Decussate tetrad type : In this two microspores lies at the right angle of other microspores.
E.g., Magnolia
- T-Shaped tetrad : Two microspores lie longitudinally and two microspores lie at transversely in this type of tetrad.
E.g., Aristolochia & Butomopsis.
- Linear tetrad : In this tetrad all four pollens arranged in linear order. E.g., Halophylla, Halophila.
- All the above type of tetrads are found in Aristolochia elegans.
STRUCTURE OF MICROSPORE OR POLLEN GRAIN
- Pollen grain is the first cell of a male gametophyte.
- Pollen grain is surrounded by two distinct layers.
The outer layer (wall) is thick, rigid and ornamented, called exine. This layer is formed by cutin and sporopollenin. Sporopollenin is highly resistant material. It is non-biodegradable. Sporopollenin is absent in the pollen grains of Zostera.
NOTES
Due to the presence of sporopollenin, fossils of pollen grains are always found in good condition. The presence of fossils of pollen grains can forecast the presence of natural resources like petroleum, coals etc. in the earth.
- Tectum provides a characteristic sculpturing or designs over the surface of pollen grains. The internal layer is thin, soft and elastic in nature. It is called intine. It is made up at pectin and cellulose or pecto-cellulose.
- Usually, at few places on pollen grain, outer surface exine is absent or present in the form of thin layer. These thin places are called germ pore. The intine comes out through any one germ pore during the germination of pollen grain in the form of pollen tube.
- The number of germ pore, structure and ornamentation of exine is a significant feature of taxonomy.
- A detailed study of pollen grains is called palynology.
- Three colpus type (slit type) of germ pore are present in pollen grain of capsella. This type of pollen grain are called tricolpate. Only one germ pore is present in monocots hence, pollen grain is called monocolpate.
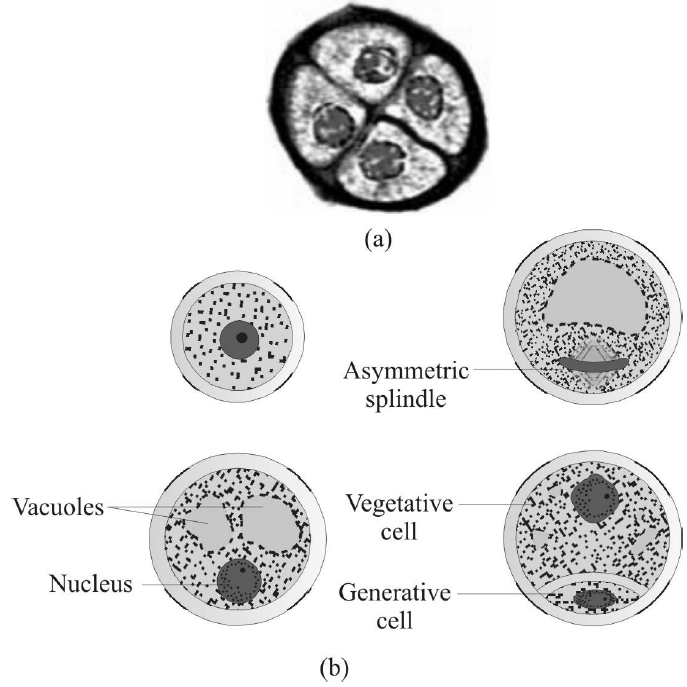
Fig. : (a) Enlarged view of a pollen grain tetrad
(b) Stages of a microspore maturing into a pollen grain
- Pollen grains of those plants in which pollination takes place by insects, have oily thick & sticky layer around the pollen grain. It is called pollen-kit which is composed of lipids and carotenoids.
- Function of pollen kit are –
- This oily layer protects the pollen grain from the harmful ultraviolet rays.
- Its sticky surface helps to attach with the insects.
- Its yellow colour attracts the insects. Pollen kit is present on the pollens of Capsella.
FACTS ABOUT TYPES OF POLLEN GRAINS
- In Asclepiadaceae (Calotropis) and Orchidaceae family, all the pollen grains join together to form "Pollinium".
- More than four pollen grains are found in tetrad called "polyspory" e.g., Cuscuta.
- In some plants, four pollens of tetrad join together permanently and are called "compound pollens", e.g., Drosera, Typha, Drimys and Elodea.
- Pollinium of Calotropis is called "Translator apparatus".
- Compound pollens of Mimosaceae (Mimosa) family contains 6-8 or 64 pollen grains stick together to form a small unit that is called "Massullae". E.g., Mimosa and Neottia plants.
- Pollen grains of some plants present in air cause allergy are called "aero allergens" E.g., Chenopodium, Parthenium, Sorghum, and Amaranthus. ("Hay fever" is caused by pollens of Ambrosia.)
- In Cyperaceae family, only one pollen grain is formed from pollen mother cell. E.g.,Cyperus.
- Largest pollen is of Mirabilis.
- Smallest pollen is of Myosotis.
- Longest pollen is of Zostera (Filliform pollen).
- Eight nucleated embryo sac type of pollen is found in Hyacinthus. [This type of pollen grain discovered by Nemec. So it is called Nemec phenomenon].
- Dumble shaped pollens are found in Vinca herbacea.
DEHISCENCE OF ANTHER
- During the maturation of anther, various changes take place in walls of anther.
- In the beginning, middle layer degenerates due to absorption of food by tapetum.
- When the micropores are formed inside the pollen sacs, at the same time ubisch bodies are formed in cells of tapetum, then after it degenerates.
In this way, in a mature anther only two layers-epidermis and endothecium are present in the form of outer covering.
- The sterile tissues are present between both the pollen sacs. Anther lobe degenerate. So both pollen sacs of each anther lobe fuse together to form single pollen sac. Therefore, in the T.S. of mature anther only two pollen sacs are present.
- Dehiscence of anther takes place during the dry season. Due to the hygroscopic nature of endothecium, loss of water takes place from the cell of endothecium.
- Walls of endothelial cells try to contract due to the loss of water but inner and radial walls do not contract due to the presence of fibrous thickening whereas outer thin walled cells of endothecium contract and become concave or incurved.
- Incurving of outer walls exert pulling force or tension over the entire surface of anther. Due to tension, thin walled stromal cells breaks off and dehiscence of anther takes place and pollen grains are present in pollen sacs released into the atmosphere.
- Dehiscence of anther in angiosperms is either longitudinal or apical pore, or transverse or valvular type. Dehiscence of anther of Capsella is longitudinal.
MICRO-GAMETOGENESIS OR DEVELOPMENT OF MALE GAMETOPHYTE
- In flowering plants, pollen grain or microspore is considered as the first cell of male gametophyte.
- Germination or development of pollen grain takes place before dehiscence of anther is called precocious development.
- Development of pollen also takes place inside pollen sac of anther is called In-situ development.
- Pre-pollination and development :
In the beginning of the process, only nucleus of pollen grain is divided by unequal mitotic division, resulting in the formation of two unequal size of nucleus. Small nucleus present near the wall is called generative nucleus and large nucleus present inside the cytoplasm is called tube or vegetative nucleus.
- Both the nucleus surrounded by cytoplasm becomes dense, followed by unequal cytokinesis, resulting into the formation of two unequal size of the cells.
- Larger cell in which large nucleus is present is known as vegetative cell and smaller cell in which small nucleus is present is called generative cell.
- Now pollen grains come in bicelled and binucleated stage. In angiosperms, pollination of pollen grains take place in bicelled and binucleated stage. The development of gametophyte take place inside the pollen grain and is also known as endosporic development.
This stage of pollen grain is called immature or partially developed male gametophyte.
- Post pollination development :
- Rest of the further development of pollen grain [Immature male, gametophyte] takes place on the stigma of carpel after pollination.
- Pollens absorb moisture and sugar content from the stigma. Due to this volume of internal content of cytoplasm increases. It exerts pressure on the both outer layers. Because of this pressure intine comes out through anyone germ pore in the form of tube like structure called pollen tube.
First of all vegetative nucleus enter into the pollen tube and assumes terminal position.
This spindle shaped generative cell now enter into the pollen tube.
- Inside the pollen tube, generative cell divides mitotically and form a two non motile male gametes.
- Now, male gametophyte comes in three celled structure in which one vegetative cell and two male gametes are present.
This three celled stage represents the mature male gametophyte of angiosperm [Capsella also]. Male gametophyte is highly reduced and completely depends on sporophyte.
- Longest pollen tube is found in Zea mays.
FEMALE REPRODUCTIVE ORGAN - GYNOECIUM
- Gynoecium is the female reproductive organ. The free unit of gynoecium is called pistil or carpel.
- Carpel is also known as megasporophyll.
- The carpel is differentiated into three distinct region –
(i) Stigma (ii) Style (iii) Ovary

Fig. : A dissected flower of Hibiscus showing pistil
(other floral parts have been removed)
(other floral parts have been removed)
- The free end of the carpel which receives pollen grains is called stigma. A long, narrow tubular structure is present in between the stigma and ovary called style. The basal swollen part of the carpel is called ovary. The ovules is also known as megasporongia which are borne on a cushion-like tissue called placenta in the ovary. One or more than one ovules are present inside the ovary.
- The gynoecium of the Capsella is bicarpellary, syncarpous, unilocular and superior. It becomes bilocular due to the formation of false septum or replum at maturity.
STRUCTURE OF OVULE OR MEGASPORANGIUM
- A ridge, or stalk-like outgrowth is formed from the placenta of the ovary on which body of ovules are present.
- Each ovule attached to the placenta by means of a thin stalk called funicle or funiculus/Funiculum.
- The point of attachment of the funicle with the ovule is called hilum.
- The main region of the ovule is composed by mass of parenchymatous cells called nucellus. Nucellus is the main part of ovule. The nucellus is covered by one or two coats called integuments.
- A place from where funicle and integuments arise is called chalaza.
Integument is absent just opposite to the chalaza, so that a narrow passage (pore) is formed which is called micropyle.
- Micropyle in bitegmic ovule has two parts -
- the outer region which is surrounded by outer integument and is called exostome.
- the inner part of micropyle which is surrounded by inner integument called endostome.
- In most of the angiosperm, entire part of the nucellus is utilized by developing embryo sac but in some of the angiosperm some part of the nucellus remain inside the ovules. That part of the nucellus present inside the seed in the form of a thin layer is known as perisperm. Perisperm is commonly found in Piperaceae (Piper nigrum) and Zingiberaceae Families (Turmeric, Ginger)
- Some filaments are attached with funicle (some times placental) are known as "Obturators". The function of obturators is to guide the passage of pollen tube towards the micropyle inside the ovary.
- In most of the ovule, funicle is attached to the main body of ovule for some distance (at lateral side) to form a ridge like structure known as Raphe.
- Vascular tissues are present inside the funiculus which supply food material from the placenta to the body of ovule.
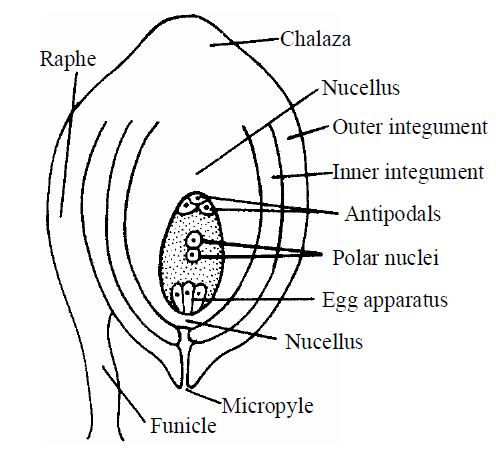
Fig. : Structure of ovule
- The placenta is the cushion-like structure to which the ovules are attached.
Some special Integuments
- Aril : It is the type of third integuments which develop from funicle at the base of the ovule. E.g., Myristica, Asphodelus and Litchi.
- Operculum : It is a stopper or break like structure which is formed on the micropyle. It is formed due to the elongation of inner integuments, E.g., Lemnaceae family (Lemna).
- Caruncle or Strophiole : It is formed due to the proliferation (out growth) of outer integuments over the micropyle. It is formed by sugary contents so helps in absorption of water during germination of seeds and dispersal of seeds by ants called myrmecochory.
E.g., Ricinus communis (Castor).
TYPES OF OVULES
ON THE BASIS OF INTEGUMENTS
- A single integumented ovule is called unitegmic ovule, e.g., members of Gamopetalae and Gymnosperm.
- Two integumented ovule is called bitegmic ovule, e.g., in most of Angiosperm [Polypetalae-Capsella and Monocots].
- The ovule in which integuments are absent is called Ategmic ovule, e.g., Olax, Liriosma, Loranthus & Santalum.
ON THE BASIS OF NUCLEUS
- Tenuinucellate : The nucellus is either less developed or present in the form of single layer. It is the most primitive type. E.g., Gamopetalae group.
- Crassinucellate : The nucellus is massive type i.e., it is made up of many layers. E.g., Polypetalae group and monocots.
- The nucellus degenerates in plants of the Compositae family and integuments becomes active to form a nucellus like tissue. This is called endothelium or integumentary tapetum. It is polyploid structure.
- The nucellus dissolves in the members of Podostomaceae family to form a nutritional cavity. This is termed as pseudo-embryo-sac.
ON THE BASIS OF RELATIONSHIP OF THE MICROPYLE, CHALAZA, AND HILUM WITH BODY OF THE OVULE AND ORIENTATION ON THE FUNICULUS
- Atropous or Orthotropous : The body of ovule is upright in position. The micropyle, chalaza and hilum lie in one straight line, so that this ovule is called straight or upright ovule. E.g., Betel, Piper, Polygonum and in Gymnosperms. It is the most primitive and most simple type of ovule of angiosperms. Raphe is absent.
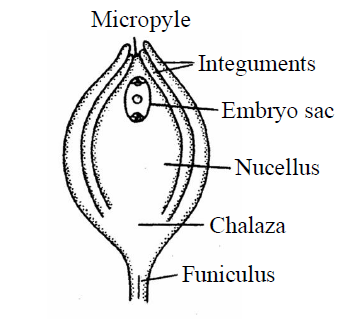
Fig. : Orthotropous
- Anatropous ovule : In this type, the body of the ovule is completely turned at 180° angle, due to unilateral growth of funiculus, so it is also called inverted ovule. The chalaza and micropyle lie in a straight line. The hilum and micropyle lie side by side very close to each other. This type of ovule is found in 80% families of angiosperms but not in Capsella. In this ovule, micropyle is facing downward condition. This is the most common type of ovule so that it is considered as a "typical ovule" of angiosperms. E.g., Members of Malvaceae, Cucurbitaceae, Solanaceae, Compositae family. It is also called resupinate ovule.
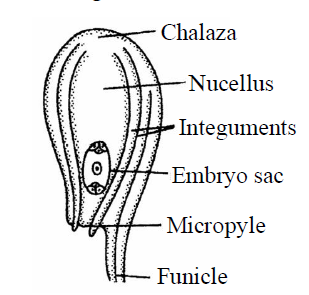
Fig. : Anatropous
- Hemitropous or Hemi-anatropous ovule : In this ovule, the body of the ovule bents on funicle at 90° angle, i.e., body of ovule is present at right angles to the funiculus. This is an intermediate type between ortho and anatropous ovules. This ovule is also called horizontal ovule because body of ovule is present in horizontal position on the funiculus. Micropyle and chalaza are present in the same line but micropyle is situated away from hilum. E.g., Ranunculus, Primula, Golphimia.
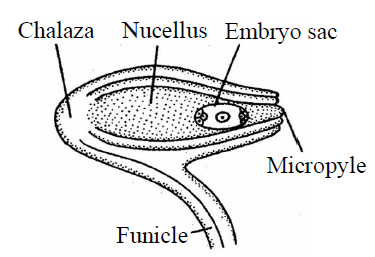
Fig. : Hemianatropous
- Campylotropous ovule : In this ovule, the body of ovule curved (Curvature is not effective) in such a way so that micropyle and chalaza are not present in straight line. The embryo sac and nucellus both are present in curved position. Micropyle comes close to the hilum. E.g., Leguminosae, Capparidaceae, Cruciferae family [Capsella]
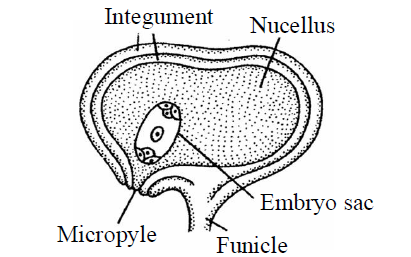
Fig. : Campylotropous
- Amphitropous ovule : In this type of ovule, curvature is more pronounced or effective in the nucellus and due to this effect of nucellus, embryo sac becomes horse shoe shaped. Micropyle comes close to the hilum. It is also called as transverse ovule. E.g., Mirabilis, Lemna and Poppy, Alisma, Butomaceae family.
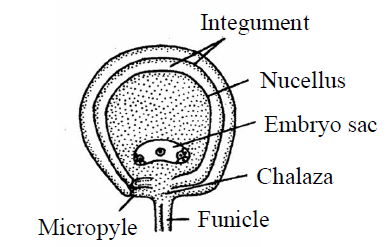
Fig. : Amphitropous
- Circinotropous ovule : In this type of ovule, body of ovule is inverted once and again turned into straight position due to the growth of funiculus so that body of ovule is present at funicle at 360°. The entire body of ovule is surrounded by funiculus. It is also known as coiled ovule. Micropyle is situated away from hilum. E.g., Cactaceae family-Opuntia.
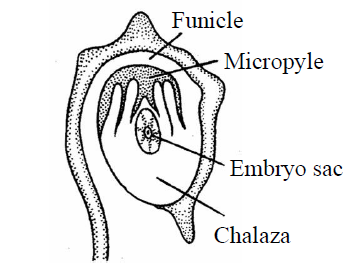
Fig. : Circinotropous
MEGASPOROGENESIS
- The process of formation of megaspore from megaspore mother cell by meiotic division is known as megasporogenesis.
- During the development of ovule, in the beginning of this process, nucellus develops from the placenta in the form of a small rounded outgrowth like structure. At this stage, all the cells of nucellus are undifferentiated and homologous and meristematic. This mass of cells is surrounded by single celled thick layer of epidermis.
A single hypodermal cell of nucellus is differentiated and increase in size. It becomes different from the rest of the cells due to the presence of distinct nucleus. It is called archesporial cell.
- Archesporium divides periclinally to form an outer primary parietal cell and inner primary sporogenous cell. Activity of primary parietal cell depends a type of plants. If plant belongs to gamopetalae then it forms tenuinucellate type ovule and if plant belongs to polypetalae then it forms crassinucellate type of ovule. The primary sporogenous cell directly act as a megaspore mother cell. It is diploid & divides meiotically to form four haploid megaspores.
The four haploid megaspores generally arranged in linear tetrad. Generally, the lowermost or chalazal megaspore remains functional. Out of tetrad of megaspores and the other three which lie towards the micropyle degenerate. This functional megaspore produces female gametophyte. In most Angiosperms [Capsella], chalazal megaspore remains functional.
DEVELOPMENT OF EMBRYO SAC OR FEMALE GAMETOPHYTE
MEGAGAMETOGENESIS
- Megaspore is the first cell of the female gametophyte.
- This megaspore grows in size and obtains nutrition from the nucellus. The nucleus of megaspore divides mitotically to form two nuclei. Each nucleus moves towards the opposite pole and reached at their respective poles. Both the nuclei that lie at poles divide twice mitotically. Resulting, four-four nuclei are formed at each poles [Total 8-nuclei].
- Out of the four, one-one nucleus migrates from both poles [one nucleus from chalazal side and one nucleus from micropylar side] towards the centre. They are known as polar nuclei. Both polar nuclei are present in the centre.
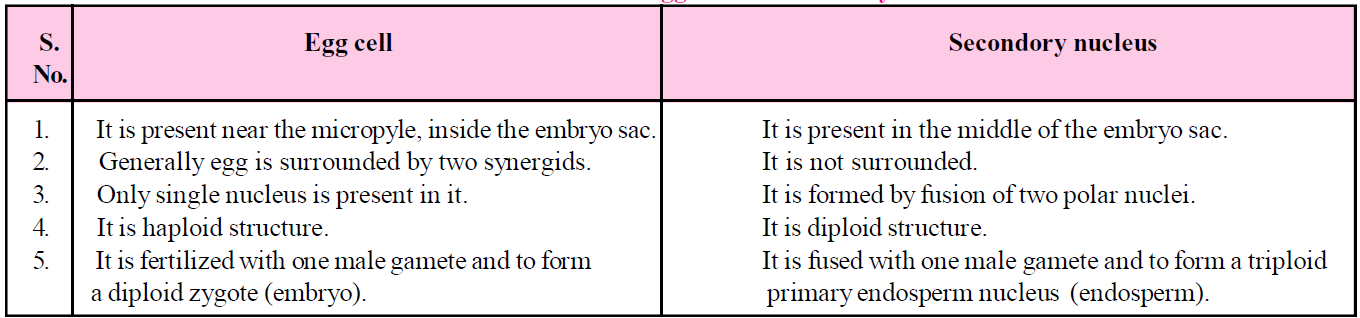
Remaining three-three nuclei at each pole is surrounded by cytoplasm to form cells as a result of cytokinesis. Three cells are formed towards the micropyle in which one cell is large and more distinct out of three cells. This is called egg cell and remaining two smaller cells are known as synergids. These three micropylar cells is collectively known as egg apparatus. [1 Egg cell + 2 Synergids]
The three cells formed toward the chalaza are called antipodal cells. Both the polar nuclei are present in the central cell. But just before the process of fertilization they unite or fuse together in the centre to form secondary nucleus. It is diploid in nature [2n] and one in number.
Therefore, seven cells and eight nucleated structure is formed. This eight nucleated and seven celled structure is called female gametophyte or embryo sac of angiosperms. This type of embryo sac is known as "polygonum type" because it is discovered by Strasburger in Polygonum plant.
- Polygonum type embryo sac is most common type in angiosperms [Capsella]. Polygonum type of embryo sac develops from single megaspore so it is also known as monosporic embryo sac.
- Finger like processes are produced from the outer wall of the synergids and are known as filiform apparatus. With the help of these structures, synergids absorb food from the nucellus and transfer to the embryo sac. Filiform apparatus is less developed in antipodal cells. Filiform also secrete chemicals which attract the pollen tube.
- In some plants, barriers are present either above or below the female gametophyte. These barriers are made up of thick walled cells of nucellus. They prevent the movement of embryo sac towards the chalaza or micropyle. The barrier which is present towards the chalaza is called "hypostase". E.g., Umbelliferae family and Zostera and Crozophora plants.
- The barrier which is present toward the micropyle is called "epistase", e.g., Costalia and Costus.
TYPE OF EMBRYO SACS
MONOSPORIC EMBRYO SAC
- It is of two types – polygonum and b.
- Polygonum type is eight nucleated and seven celled embryo sac.
- Exceptionally oenothera type is four nucleated in which only one nucleus is in a central cell and three nucleus in egg apparatus. Antipodal cells are absent. [Micropylar megaspore become functional].
BISPORIC EMBRYOSAC
- It is formed by two megaspores. It means it develops from two nuclear megaspores.
- It is of two types –
- Allium type - Eight nucleated and seven celled [Chalaza megaspores]
- Endymion type - Eight nucleated and seven celled [Micropylar megaspores]
Tetrasporic Embryo sac
- It is formed by all four megaspore nuclei because meiosis is not accompanied by cytokinesis, so that four nuclei of megaspores are formed.
- All four nuclei are collectively known as "Coenomegaspore".
- Generally, all the four nuclear megaspores take part in the formation of the seven celled and eight nucleated embryo sac but different types of embryo sacs are also formed. Arrangement of nuclei are not definite in embryo sac in some angiosperms, so that different type of embryo sacs are formed.
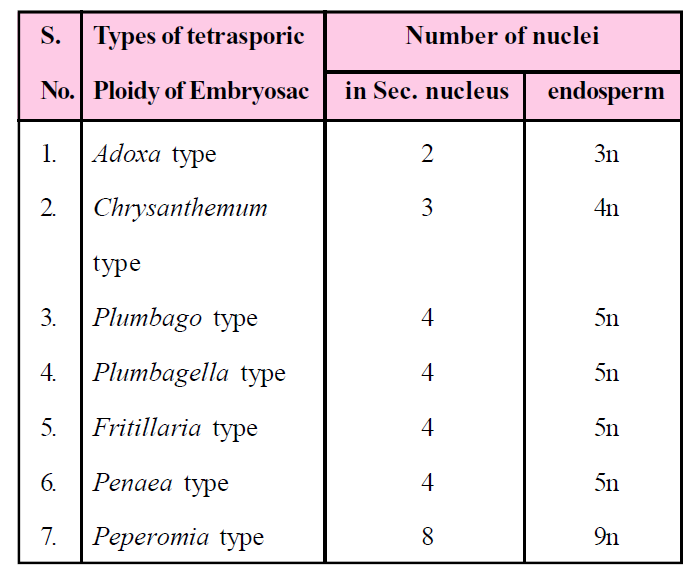
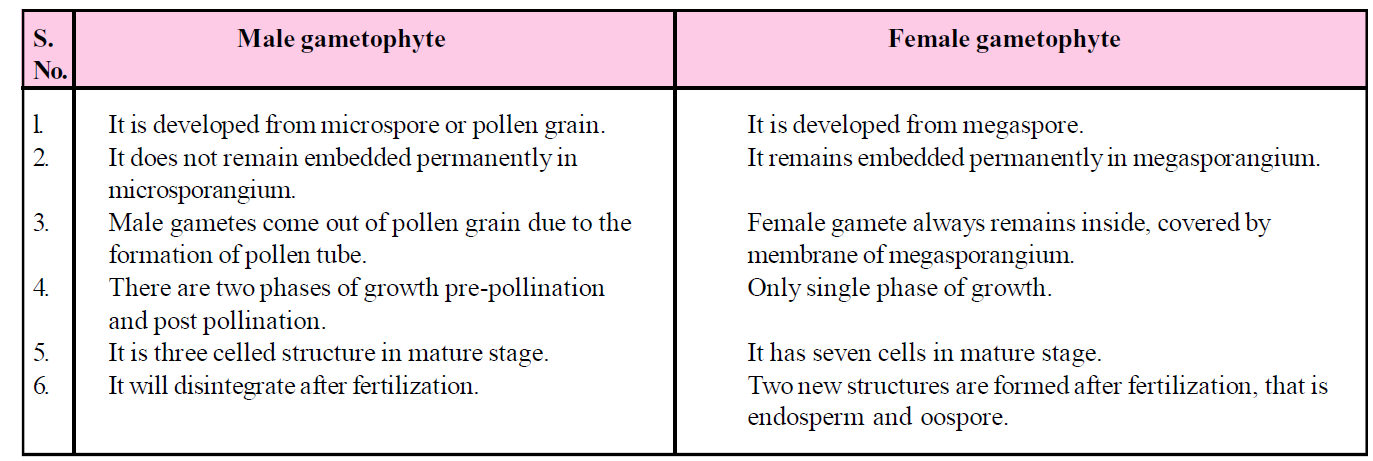
Table : Difference between male and female gametophyte
POLLINATION
- Pollination is defined as the process of transfer of pollen grains from the anther to the stigma of the same flower or of different flower of the same species.
- The process of pollination occurs only in gymnosperms and angiosperms.
- Pollination is of two types : self pollination and cross polination.
SELF POLLINATION OR AUTOGAMY
- If the pollen grain are transferred from an anther to the stigma of the same flower, or different flowers of the same plant is called self pollination or autogamy.
ADAPTATION FOR SELF POLLINATION
- Monocliny :- Bisexuality it means flowers are bisexual.
- Homogamy:- When both the sex organs of a flower mature at the same time is called homogamy. E.g., Mirabilis, potato, sunflower, wheat, rice, etc.
- Cleistogamy : In some plants, bisexual flower are formed which never opens throughout the life. Such flowers are called cleistogamous flowers such as Commelina, Viola. Commelina plant have two types of flowers. One type of flowers are cleistogamous and another are chasmogamous flowers (In which pollination occurs in opened flower, which expose their sex organs).
A special type of cleistogamy is found in flowers of legume plant. The sex organs are closed in a structure which is formed by joining of some petals. It is called keel. This keel never opens. So, only self pollination takes place in these plants.
CROSS POLLINATION OR ALLOGAMY
- When the pollen grains are transferred to the stigma of other flower of the same species is called cross pollination or allogamy. It takes place in between two different flowers.
- Cross pollination is of two types – Geitonogamy and Xenogamy.
- Geitonogamy : When pollination takes place in between the two flowers of the same plant then it is called geitonogamy. From the genetical point of view, geitonogamy is self pollination because all the flowers of the same plant are genetically identical. But ecologically, it is considered as cross pollination.
- Xenogamy : When the pollination takes place in between the two different flowers of two different plants of the same species then it is called xenogamy. This is real or true cross pollination. Genetically, as well as ecologically, it is cross pollination.
ADAPTATION FOR CROSS POLLINATION
- Dicliny : Unisexuality of the flower confirms cross-pollination. Self pollination never takes place in these flowers. It means allogamy becomes compulsory.
Examples - Morus, Palms, Date Palms, Cucumber and Cucurbita, Carica etc.
- Dichogamy : In many hermaphrodite flowers of the plants, stamens and carpels of a flower do not mature at the same time. Dichogamy is of two types–
- Protandry : The anther of a flower mature earlier than carpels, is called protandry. In most of the plants of angiosperms, cross pollination occurs only because of protandrous condition. E.g., Salvia.
- Protogyny : The carpels of the flower mature earlier than stamens. It occurs in very less no. of plants, e.g., Ficus bengalensis, Saraca indica, Ficus religiosa, Aristolochia, most of the plants of Cruciferae and Rosaceae family.
- Chasmogamy or anthesis : Opening (blooming) of the floral bud in the form of a flower is called anthesis.
- Herkogamy : In some plants, morphological barriers are formed in between the anther and stigma of the same flower, so self pollination cannot occur and pollen grains from the anther are unable to reach the stigma of the same flower. In this condition, only cross pollination is possible. E.g., Gloriosa, Calotropis and Caryophyllaceae family.
- Heterostyly : There is a difference between the length of the filaments of stamens and length of style in flowers of some plants. Some of the plants having long stamen and short style, and in some of the plants bears long style and short filament. Due to this reason, self pollination is not possible in these plants, e.g., Primrose and Linum, Primula.
- Self sterility or self incompatibility or intraspecific incompatibility (out breeding devices) : In this condition, the pollen grains of the flower cannot germinate on the stigma of the same flower. This condition is called self sterility. This is a parental (genetical) characteristic feature which is controlled by genes. Such as in Pitunia, Malva, Thea, Passiflora, Vitis, Apple (Malus).
Incompatibility involves many complex mechanisms associated with interactions of pollen and stigmatic tissues. If the incompatibility is due to the genotype of the sporophytic/stigmatic tissues, it is termed sporophytic incompatibility. On the other hand, if it is due to the genotype of the pollen, it is termed gametophytic incompatibility. This may be due to prevention of pollen germination, retardation of growth, disorientation of pollen tube or even failure of nuclear fusion. It is controlled by genes with multiple alleles (s-allele).
METHOD OF CROSS POLLINATION
- Cross pollination requires external agents for the transfer of pollen grains from one flower to another flower like air, insects, water, bird, etc.
Anemophily
- When the pollen grains are transferred from one flower to another flower through the air it is called anemophily and flower is known as anemophilous flowers. Such as cereal plants, Maize.
Anemophily is also found in all gymnosperms.
The anemophilous plants produce enormous amount of pollen grains. The pollen grains are very small, light weight and dry and their stigma is hairy or brushy and mucilagenous (Sticky).
Anemophilous flowers are neither attractive nor with fragrance. They do not have nectar glands. Anemophilous flowers are generally unisexual.
E.g., Sugarcane, Bamboo, Coconut, Cannabis, Grasses and Date palms, Typha, Oak, Hazel.
- Yellow clouds are formed in the sky during the wind pollination in Pinus. These yellow clouds are formed due to the pollen grains and are called "Sulphur Showers".
- Potamogeton and Myriophyllum are aquatic plant but it is anemophilous.
- Maximum loss of pollen grains takes place in this pollination. It is completely non directional process.
Entomophily
- The pollination that takes place with the help of insects is known as entomophily. Most of insect pollination (80%) occurs only by honey bees. They are ornamental plants.
- Ornamental plants utilize their maximum energy in this pollination and develops different types of adaptation for attraction of insects. These flowers are attractive in colour. They posses special fragrance. Nectar glands is also present.
- The pollen grains of insect pollinated flowers become sticky due to the presence of pollen kitt. The surface of stigma of flowers is rough.
Some of the plants develop special adaptations for insect pollination.
- In Mussaenda plant, advertising flag is for attraction of insects.
- Some of the flowers have attractive bracts i.e., bright and coloured i.e., Bougainvillea.
- Yucca plant has developed an obligate symbiotic relationship with an insect Pronuba moth. The pollination in Yucca takes place only by Pronuba moth. This insect lays eggs in the ovary of a flower. Life cycle of both depend on each other.
Some other mechanism of Pollination
- "Trap door mechanism" is found in species of Ficus for pollination [Blastophaga insect] because hypanthodium type of inflorescence is present.
- In Aristolochia "Fly trap mechanism" is found for pollination. This flower is known as "Pit fall flower".
- The lever mechanism or turn pipe mechanism is found in Salvia for pollination. The flowers of Salvia have bilabiate corolla tube. The connective of stamen is long. The anterior anther lobe of connective is fertile while posteriorly lobe is sterile. When the insect lands on the lower lip, the fertile lobe automatically brings down to touch the back of insect and thus depositing the pollen grains on the back of insect.
- Ophrys speculum flower is pollinated by Wasp [Colpa aurea] by means of pseudo-copulation. The appearance and odour of the flower like female wasp [Mimicry].
- In Centaurea plant, pollination takes place by "piston mechanism".
- In Rafflesia, the pollination is brought about by Carrion flies and dispersal of seeds by elephant.
- Nymphaea, Nelumbo and Alisma are also entomophilous plants while they are hydrophytes.
Hydrophily
- When the pollination brings by water is known as hydrophily. It is of two types - epihydrophily and hypohydrophily.
- Epihydrophily : When the pollination takes place on the surface of water, i.e., Vallisneria and Ruppia.
- Hypohydrophily : When the pollination takes place inside the water, i.e., Ceratophyllum, Najas and Zostera and Hydrilla. Specific gravity is found in pollen grains so remains suspended in water.
Zoophily
- When the pollination brings by animals is called zoophily. Generally in zoophillous plants, flowers are very large, attractive and maximum number of nectar glands are present (more than entomophillous).
- Zoophily is of following types-
- Ornithophily : The process of pollination takes place by birds is known as ornithophily. E.g., Sun bird or humming bird in Begnonia plant and by Honey bird in Strelitzia plant, Callistemon (Bottle brush), Bombax [Silk cotton tree,] Butea monosperma.
- Chiropterophily : If the pollination brings through the bats then it is called chiropterophily. E.g., Pteropus. These flowers are big in size, i.e., Anthocephalus kadamba, Bauhinia & Kigelia plants, Adansonia.
- Myrmecophily (Termite pollination) : This pollination brings about by Termites and Ants. E.g., Prosopis kikar, Acacia, (Mimosoideae family) and some members of Rubiaceae family etc.
- Malacophily or Malmacophily : This pollination brings about by Snails, i.e.,Aspidistra lurida and Chrysanthemum plants.
- Erythrina (Coral tree) plant is visited by Crows and Squirrels.
- In Arisaema or Cobra Plant, pollination occurs by snake called "Ophiophily".
- Night flowering plants are pollinated by moths. They are highly scented.Their flower generally white coloured.
- Orchids are pollinated by wasps/snails.
POLLEN PISTIL INTERACTION
- It is an active process involving pollen recognition followed by promotion or inhibition of the pollen.
- Pollen pistil interaction provides a unique opportunity in plants for elucidating the molecular basis of cell-cell communication as well as for understanding the operation and evolution of intra-specific and inter-specific barriers to fertilization.
EMASCULATION AND BAGGING
- Emasculation is the process of removal of stamens of flowers before they have shed pollen in order to prevent self pollination and permit cross pollination.
- Emasculated flower have to be covered with a bag of suitable size, generally made up of butter paper, to prevent contamination of its stigma with unwanted pollen. This process is called bagging.
Table : Difference between pollination and fertilization
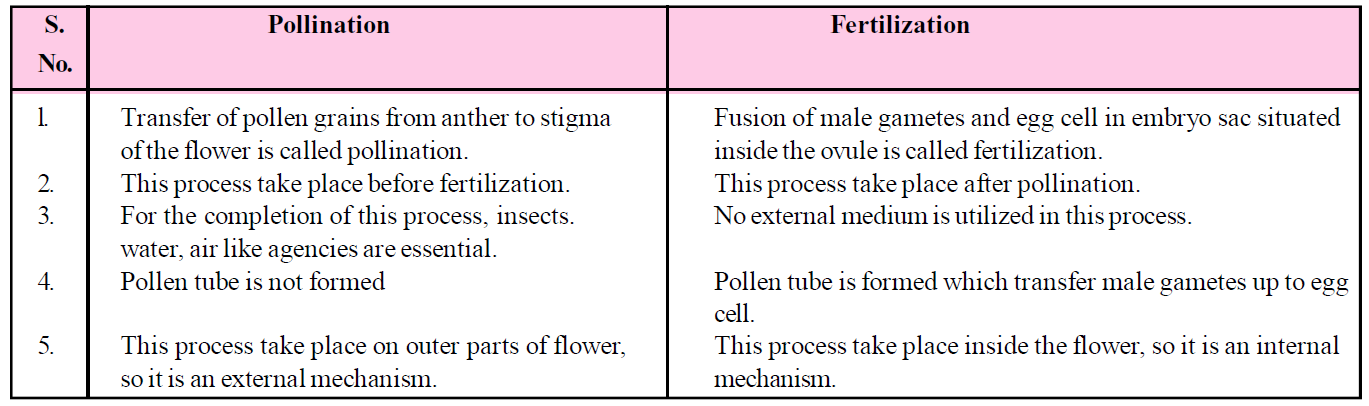
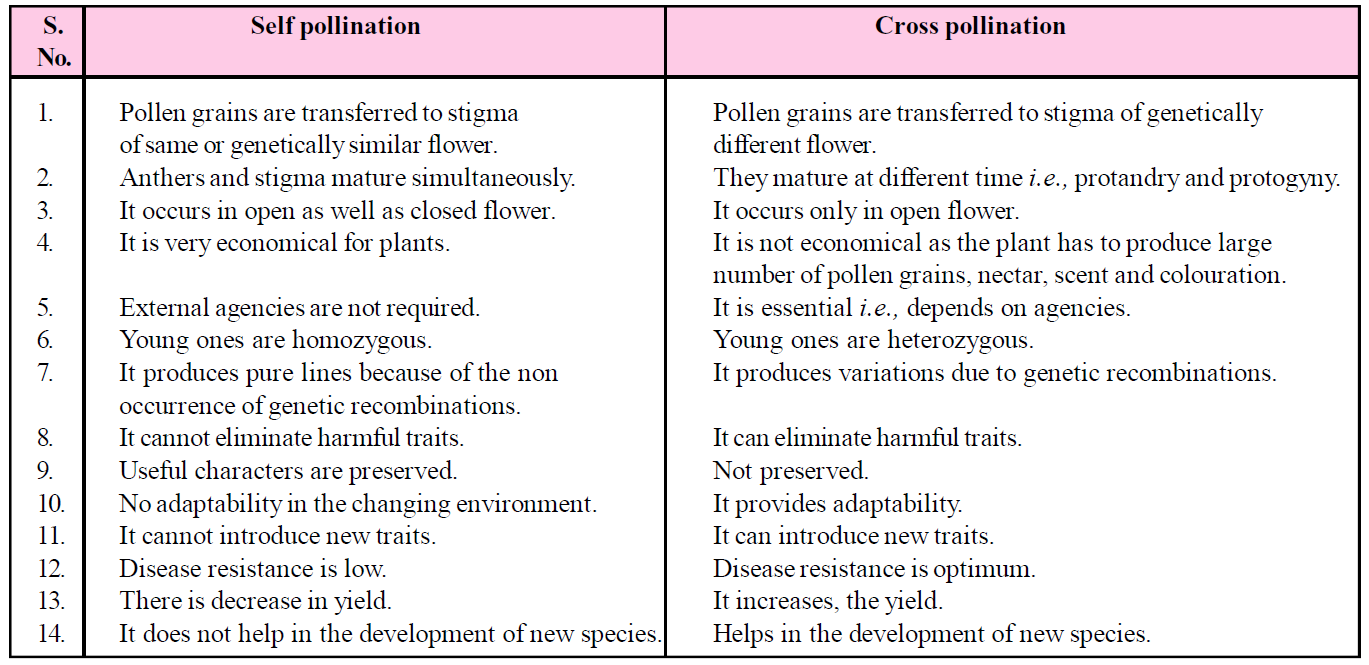
Table : Difference between Self and Cross pollination
FERTILIZATION
- The fusion of male gamete with female gamete is called fertilization.
- Fertilization was discovered by Strasburger (1884) in Monotrapa plant. This process of fertilization is completed in the 4 steps :
- Germination of pollen grain and pollen tube.
- Entry of pollen tube into the ovule.
- Entry of pollen tube into embryo sac.
- Fusion of gametes.
GERMINATION OF POLLEN GRAIN
- After pollination, pollen grains germinate on the stigma of the carpel. They absorb moisture and sugar contents from stigma and swell up.
- The intine of pollen grain grows out through the anyone germinal pore of exine, in the form of tube like outgrowth called pollen tube.
- One pollen tube develops in Capsella and most of Angiosperms is called monosiphonous condition, but more than one pollen tube develops in Malvaceae and Cucurbitaceae family and called as polysiphonous.
- When the pollen tube comes down from the stigma into the style, first of vegetative nucleus enter, into the pollen tube then it is followed by generative cell. The tube nucleus always occupies in terminal position in pollen tube. The vegetative nucleus controls the growth of the pollen tube. Meanwhile, the generative cell divide mitotically to form two male gametes. Both of the male gametes are non motile.
- Boron element and calcium ions (mainly Boron) are essential for the growth of pollen tube and best temperature for growth of pollen tubes is 20-30°C. Pollen tube shows apical growth and chemotropic movement.
ENTRY OF POLLEN TUBE INTO THE OVULE
- The pollen tube enters in the ovary at the time when ovule becomes mature.
- Inside the ovary, obturators guides the passage of pollen tube towards the micropyle.
- A mature ovule in which the embryo sac also matured, has three paths for the entry of pollen tube:
- Porogamy : In this, pollen tube enters into the ovule through the micropyle. It is found in most of Angiospenns [Capsella].
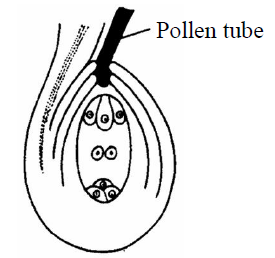
Fig. : Porogamy
- Chalazogamy : In this method, the pollen tube enter into the ovule through the chalaza. This method is discovered in Casuarina by Treub [1891], e.g., Betula and Juglans (walnut).
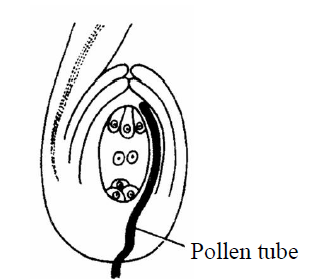
Fig. : Chalazogamy
- Mesogamy : In this method, pollen tube enter into the ovule either through integuments (Cucurbita) or through the funiculus (Pistacia and Populus).
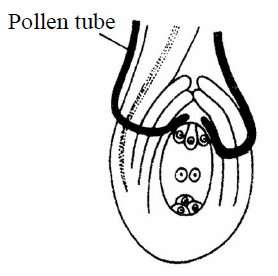
Fig. : Mesogamy
ENTRY OF POLLEN TUBE INTO EMBRYO SAC
- Pollen tube can enter into the ovule through any passage but inside embryo sac, it enters only through the egg apparatus. After the entrance inside the ovule, it grows towards the egg apparatus because synergid cells secrete the chemical (hormones) which attracts the growth of pollen tube. It means pollen tube shows chemotropic movement in ovule.
- Any one synergid starts degenerating when the pollen tube comes near egg apparatus. The pollen tube enters into the embryo sac through the degenerating synergids.
- When tip of the pollen tube enters into the embryo sac, vegetative nucleus degenerates. The tip of the pollen tube swells and burst (endosmosis) after reaching inside the embryo sac. The pollen tube releases all contents including both male gametes inside the central cell of the embryo sac.
- Two dark granules appears in the region of degenerating synergids. These are known as X-bodies. They are two in number and both X-bodies are formed by the degenerating nucleus of tube cell and synergids.
FUSION OF GAMETES
- Before or after the entrance of pollen tube into the embryo sac both polar nuclei of the central cell fused together to form a diploid nucleus. It is known as secondary nucleus or definitive nucleus.
- Out of two, one male gamete fertilized with egg cell and form a diploid zygote. This fusion is known as syngamy. This is true mechanism of fertilization process.
The second male gamete fused with diploid secondary nucleus which is formed by the fusion of two polar nuclei. This fusion is known as triple fusion resulting in a formation of triploid (3n) structure. It is called primary endosperm nucleus.
- Fertilization takes place twice at a time in angiosperm called double fertilization.
Double fertilization was discovered by "Nawaschin" in Lilium and Fritillaria plants.
- Double fertilization and triple fusion is the specific or universal characteristic of Angiosperm. There are five nuclei and three gametes participate in double fertilization.
- A zygote is formed by true fertilization (syngamy) and develops into an embryo. Triploid primary endosperm nucleus occurrence of supernumerary male gametes is called "polyspermy." is formed by triple fusion that develops into the endosperm which is used as nutrition for the growing embryo.
- All the remaining cells of embryo sac like antipodal cells, synergids degenerate excluding zygote and primary endosperm nucleus after fertilization. At this time, zygote obtains food from degenerating synergids and antipodal cells.
- The fertilization in which non motile gametes are carried to female gamete through pollen tube is known as "siphonogamy".
- Entry of more than one pollen tube into the ovule leading to occurrence of supernumerary male gametes is called "polyspermy."
Table : Difference between egg cell and secondary nudeus
DEVELOPMENT OF ENDOSPERM
- Endosperm which stored food materials develops from the primary endosperm nucleus after fertilization . This food material is utilized by the embryo during the early development and then after at the time of seed germination. Food is present in the form of starch in endosperm.
- Endosperm is a nutritive tissue which provide nourishment to the embryo in a seed plant. It is generally triploid.
- In gymnosperm, endosperm is formed before fertilization & is always haploid.
TYPES OF ENDOSPERMS
- The endosperm is of three types on the basis of development – nuclear, cellular and helobial.
NUCLEAR ENDOSPERM
- Nuclear endosperm is mostly found in Dicotyledon (Polypetalae) and it is also present in Capsella.
- This type of endosperm develops by free nuclear divisions of nucleus of primary endosperm nucleus, resulting in multinucleated endosperm. Later on, cytokinesis takes place, so that multicellular endosperm is formed.
- It is the most common endosperm in Angiosperms.
- The milky fluid, found in green coconut, is an example of nuclear endosperm, which is called liquid syncytium.
CELLULAR ENDOSPERM
- This type of endosperm is found in gamopetalae group, e.g., Petunia, Datura.
- During the development, each division of primary endosperm nucleus is followed by cytokinesis, so that endosperm remains cellular from the beginning.
HELOBIAL ENDOSPERM
- During the development of this type of endosperm, first division of primary endosperm nucleus is followed by unequal cytokinesis so that two unequal sized cells are formed (Cell towards the micropyle is large). Now free nuclear divisions takes place in each cell, and becomes multinucleated. Eventually cytokinesis takes place at a time so that it is changed into a cellular endosperm. This type of endosperm is found in monocots. It is an intermediate type of endosperm.
NOTES
- After the observation of the first two divisions of primary endosperm nucleus, endosperm can be identified whether it would be nuclear, or cellular endosperm.
- Endosperm is absent in some angiosperms like Orchidaceae, Podostomaceae and Trapaceae.
- Exceptionally, some of the plants have diploid endosperm instead of triploid such as in Oenothera.
- Maize and Tomato have mosaic endosperm in which patches of different colours are present.
- The endosperm in Betelnut is rough surfaced. It is known as "ruminate endosperm".
- In coconut, the drinking portion is nuclear endosperm and edible portion is cellular endosperm.
DEVELOPMENT OF EMBRYO
- The process of development of mature embryo from zygote or oospore is called embryogeny.
- Zygote starts dividing to produce embryo, together with the development of endosperm.
DEVELOPMENT OF EMBRYO IN DICOTYLEDON
- In angiosperm, zygote undergoes a resting phase. When the endosperm is formed, development of zygote starts. In the beginning, it absorb food from the endosperm and increases in size then later a layer is secreted by itself. Now it is called oospore.
- The first division of oospore is transverse. As a result, two cells are formed. The one cell lies towards micropyle and called basal cell or suspensor cell. The other cell is formed towards the chalaza and is called apical cell or terminal cell or embryonal cell.
- The basal cell and embryonal cell divide simultaneously.
- The basal cell divides transversely and apical cell divides vertically and resulting in two suspensor cells and two embryonal cells. This stage is made up of four cells which are arranged in 'T' shaped structure. Embryonal cells divide vertically to form four embryonal cells. This is the quadrant stage of embryo.
- The two suspensor cells divided by transverse divisions forming a 6-10 celled long filament like structure, called as suspensor. The main function of suspensor is to push the developing embryo into food laden endosperm to provide nutrition.
- The micropylar cell of the suspensor swells up. This cell of suspensor is known as haustorial cell.
- The cell of suspensor lies near the embryonal cells and is called hypophysis. This cell combines with radicle to form the apex of root (Root cap).
- These four celled quadrant embryo further divide transversely to produce eight celled stage of embryo (called octant stage). The eight cells of octant are arranged in two tiers.
- The four cells of the octant embryo lies near the hypophysis is known as hypobasal cells and four cells present towards the chalaza is termed epibasal cells.
The hypobasal cells gives rise to radicle and hypocotyl and epibasal cells gives rise to two cotyledons and plumule of the embryo.
- All the cells of octant divides by periclinal division so that a 16 celled globular embryo is formed (proembryo).
Due to fast division of embryonal cells of globular embryo, a heart shaped embryo is formed. All the cells of this embryo are meristematic.
- Due to the fast growth in two lobes of heart shaped embryo, they develop into two cotyledons. Both the growing cotyledons turn in downwards direction due to the curved position of body of ovule of Capsella.
- The tissues are present below the joining place of both. The cotyledons are responsible to form plumule and behind it epicotyl is formed.
- The tissues present opposite to the plumule near the hypophysis give rise to radicle. The apex root (root cap) is formed by hypophysis. This curved position of the embryo is called torpedo or chordate stage.
- An axis is present between plumule and radicle and is called embryonal axis. It is also called tigellum (main embryonal axis).
- Both the cotyledons are present at lateral position of embryonal axis and plumule is formed in terminal position in Dicotyledon embryo.
- This type of development of embryo is known as Crucifer type or Onagrad type. It is the most common type of development in Dicots.
- Crucifer type of development is also found in Capsella, so that it is considered as typical angiosperm for the study of embryonic development of Angiosperms.
- When an embryo is formed, suspensor dries and degenerates so it is known as meroblastic development.
- Ovule modified into seed in which testa is formed by outer integument and tegmen is formed by inner integument.
- Only micropyle of ovule remains unchanged and also present in seed.
- Entire ovary is modified into fruit. This fruit is formed by fertilized ovary so that it is called true fruit.
- In some of the angiosperms, fruit is formed from the ovary without fertilization known as parthenocarpic fruit.
- In some fruit, parthenocarpy is senseless (If edible part is endosperm or seed).
DEVELOPMENT OF EMBRYO IN MONOCOTYLEDON
- The Lilium type of embryonic development is found in monocotyledons.
The first division is transverse division in Oospore. As a result two cells are formed– the upper cell chalazal is called embryonal cell and lower micropylar cell is termed as basal cell. The basal cell does not divide further and later on it increases in size and form single celled vesicular suspensor.
- Only embryonal cell divides transversely in which terminal cell is called cotyledon cell and lower (middle) cell is known as embryonal axis cell.
- A transverse division takes place in embryonal axis cell to gives rise two cells.
The one cell out of two, gives rise to plumule initial and another gives rise to radicle initial.
- The plumule initial divides to form a plumule of the embryo. Radicle initially divides to form radicle. In this, both the initials are responsible to form embryo in lateral position.
- An apical cotyledon is formed by the continuous divisions of cotyledon cell. This apical cotyledon is known as scutellum.
- The plumule and radicle is covered by cap like hard protective structures. The covering of plumule is called coleoptile and covering of radicle is known as coleorhiza. These structures are broken during the germination The plumule lies in lateral position and cotyledon in terminal position in monocotyledons.
Table : Difference between Embryo and Endosperm
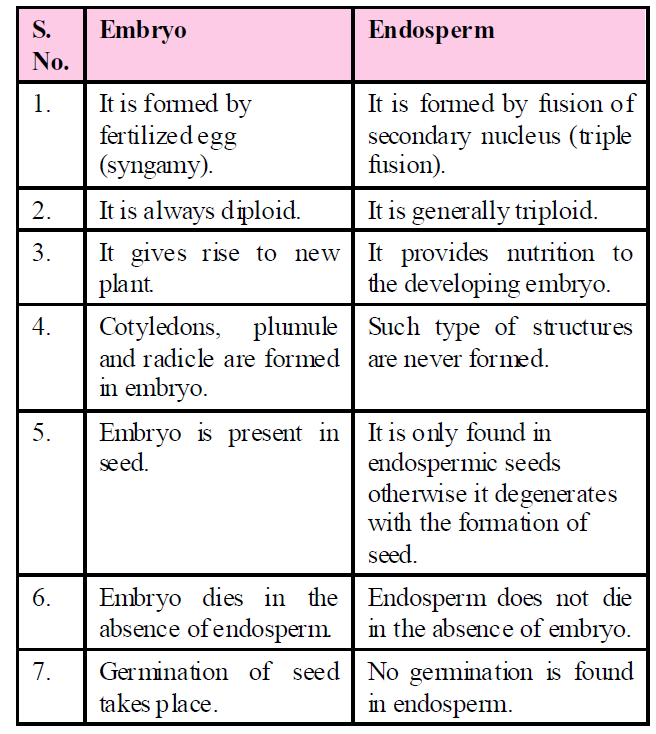
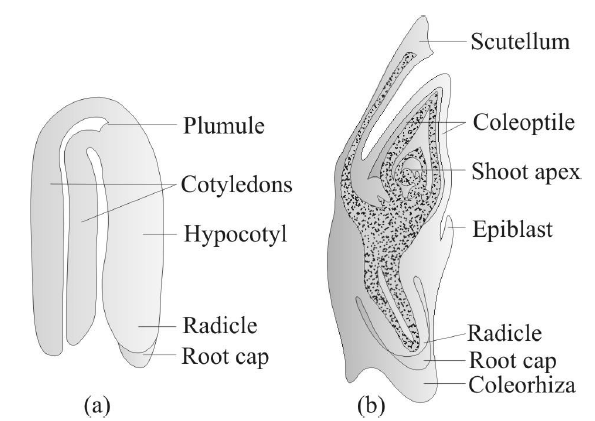
Fig. : (a) A typical dicot embryo; (b) L. S. of an embryo of grass
STRUCTURE OF SEED
- The whole plant exists in a seed. In angiosperm, after fertilization, egg cell changes into embryo, ovary changes into fruit and ovule changes into seed.
- Morphologically, ripened ovule is known as seed. In other words, seed is a mature, integumented megasporangium.
- Seeds are characteristic of spermatophytes (gymnosperms and angiosperms).
- Seed is a dormant structure containing protective coverings, reserve foods and embryo (2n).
SEED COAT
- Outer protective covering of the seed is called seed coat, which develops from integuments of ovule.
In seeds, developing from bitegmic ovules, there are two distinct layers in seed coat-the outer layer is thick, hard and leathery (developing from outer integument), called testa, whereas inner layer is thin and papery or membranous (developing from inner integument), called tegmen. In seeds developing from unitegmic ovules, there is single layered seed coat. The seed coat performs usual protective function.
- The seed is attached to the fruit wall or pericarp by means of a stalk called funicle or funiculus.
The point of attachment of funiculus to the body of mature seed is called hilum.
A small opening or pore called micropyle is present just near the hilum, which is the way of entry of water into the seed. As most of the ovules in angiosperms are inverted or anatropous, so a ridge called raphe is formed.
EMBRYO
- Embryo is the most important part of the seed, which represents tiny future plant. The origin of embryo takes place from fertilized egg (zygote).
- The embryo is having an embryonal axis or main axis called tigellum, to which one or two cotyledons (seed leaves) are attached, depending upon whether the seed is monocot or dicot.
- The portion of embryonal axis or tigellum below the point of attachment of cotyledons, is called hypocotyl, which bears radicle or future root at its tip. Similarly, portion of embryonal axis or tigellum above the point of attachment of cotyledons is called epicotyl, which bears plumule (future shoot) at its tip.
COTYLEDONS
- The cotyledons of the embryo are simple structures, generally thick & swollen due to storage of food reserves (as in legumes). In some seeds (e.g., legumes), reserve food is stored in cotyledons, whereas in others (e.g., cereals), there is special nutritive tissue called endosperm.
The reserve food materials in seeds may be carbohydrate (e.g., wheat, rice) or proteins (legumes) or fats (e.g., castor, peanut, sunflower, etc.)
- Sometime, some part of nucellus remains unused which is present in the form of thin layer around the endosperm is called perisperm. E.g., Betel nut, Black pepper, Castor, etc.
- In Monocot embryo, there is a single cotyledon called scutellum. In some grasses - wheat, on opposite side of scutellum is a tongue shaped outgrowth remains of second cotyledon is present called epiblast.
- In Dicot albuminous seed-castor (Ricinus communis), there is a specific outgrowth called caruncle or strophiole, present over micropyle. It is formed by proliferation of cells of outer integument at tip. Caruncle is somewhat spongy and helps in absorption of water during germination of seed.
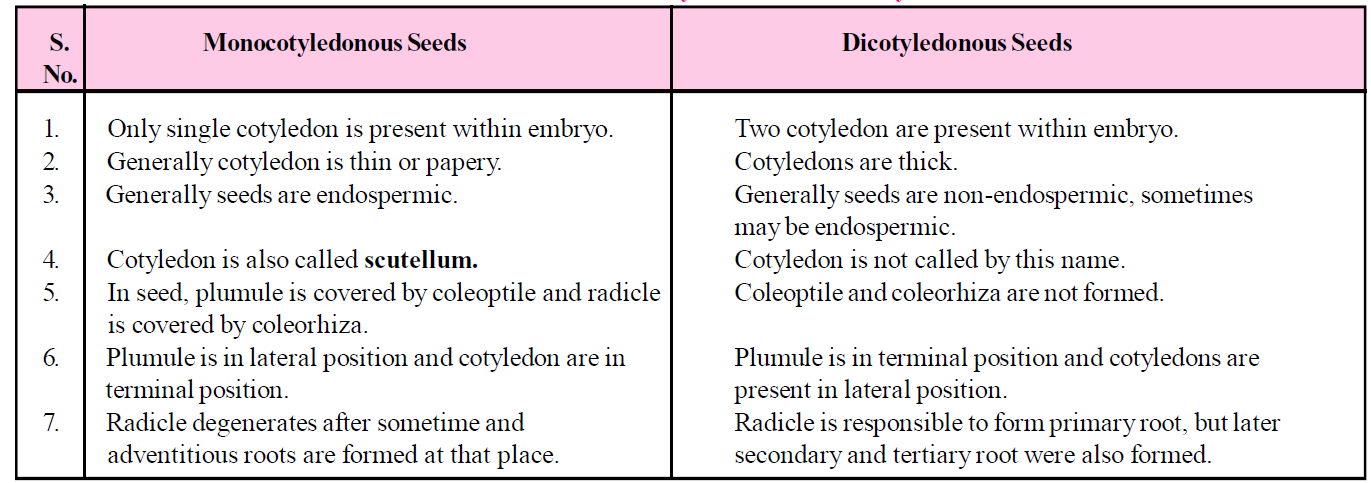
Table : Difference between Monocotyledonous and Dicotyledonous seed
TYPES OF SEEDS
On the basis of presence or absence of endosperms in seeds, it is of two types- endospermic or albuminous seeds and non-endospermic or ex-albuminous seeds.
ENDOSPERMIC OR ALBUMINOUS SEED
- In this type of seeds, food is stored in endosperm. Such seeds are called endospermic seed or albuminous seed.
- The endospermic tissue in these seeds utilize during the germination of seed and their cotyledons are thin and membranous e.g., all monocot seeds, (wheat, rice, and maize). Castor [dicot] is also endospermic.
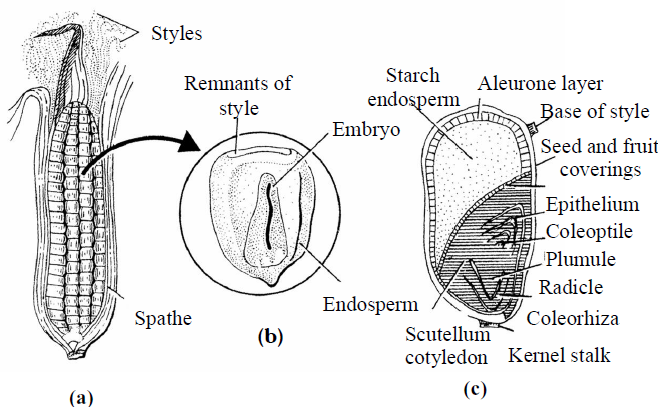
Fig. : Structure of Maize grain (a) Entire seed;(b) Grain in L.S. (c) Grain in T.S.
NON ENDOSPERMIC OR EX-ALBUMINOUS SEED
- Such type of seeds do not have an endosperm at maturity, therefore are called non endospermic e.g., albuminous seeds.
- The endospermic tissues are absorbed during the development of embryo. The absorbed food materials from the endosperm is stored in cotyledons that why they become so large and fleshy, e.g., Capsella and all dicotyledons. But Castor seed is endospermic.
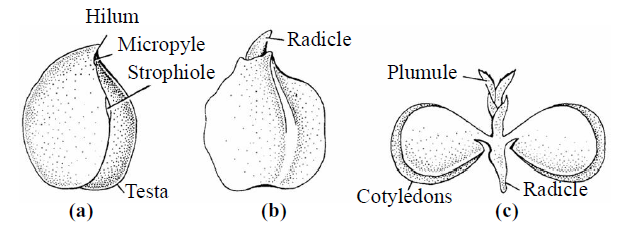
Fig. : Structure of Gram seed
Endospermic dicot seeds: E.g., Castor, Papaya, Cotton.
Non-endospermic dicot seeds: E.g., Gram, Bean, Pea, Cucumber ,Tamarind.
Endospermic monocot seeds: E.g., Maize, Rice, Wheat and Coconut.
Non-endospermic monocot seeds: E.g., Pothos (money plant), Vallisneria, Alisma, Amorphophallus.
GERMINATION OF SEED
- Seed is fertilized and ripened ovule which contains a dormant embryo, reserve food for its future development & protective covering.
- The formation of seedlings from dormant embryo of seed after resumption of its active growth, is called germination.
- Rate of respiration is increased in germinating seeds.
- Germination is basically of two types (epigeal and hypogeal) depending upon behaviour of cotyledons but viviparous germination is also found.
EPIGEAL GERMINATION
- Due to hypocotyl growth or elongation, cotyledons are pushed out of soil. This is called epigeal germination.
- This type of germination occurs in cotton, papaya, castor, onion, cucurbits, tamarind, french beans, mustard, etc.
- In some cases, the above ground cotyledons become green leaf like (cotyledonary leaves) and perform photosynthetic function till the seedling assumes independency (e.g., castor, cotton, onion, papaya, etc.). In other plants, cotyledons do not assume leaf like shape and fall off (e.g., French bean and Tamarind).
HYPOGEAL GERMINATION
- Due to first epicotyl elongation, plumule comes out of the ground and cotyledons remain underground. This is called hypogeal germination.
- This type of germination occurs in most of the monocotyledons and few dicotyledons, e.g., maize, rice, wheat, coconut, gram, pea, peanut and mango, etc.
VIVIPARY GERMINATION
- Vivipary is a special type of seed germination of Mangrove vegetation, found in muddy, saline conditions, e.g., Rhizophora, Avicennia, Ceriops, Breguira, Sonneratia. etc.
- In this, there is no resting period of embryo and germination occurs inside the fruit, while it is attached to the parent plant, i.e., "in situ germination". The seedling is separated in the mud with the help of lateral roots developing from basal end of radicle. This is called viviparous germination or vivipary.
Table : Difference between Epigeal and Hypogeal germination

DORMANCY OF SEED
- The presence of dormancy in a seed is the most important characteristic feature. Because of this character, seeds remain viable for many years. The seeds are dispersed at very far places through water, air or insects. Most of the seeds are unable to germinate just after dispersal. They germinate after sometime.
- The time between the maturation and germination of seed is known as "dormancy period". The state of inhibited germination as a result of internal causes is usually called 'dormancy'.
- This seed dormancy is of considerable advantage to the plant which helps in adverse environmental conditions. The embryo remains inactive in this period and all the growth processes suspended temporarily reasons of seed dormancy.
THREE MAIN BASIC REASONS OF DORMANCY OF SEED
IMPERMEABILITY OF SEED COAT
- The seed coats of many species of Leguminosae and Convolulaceae families are completely impermeable to moisture (water) and oxygen at the time of their maturity. Seed coats in these are thick and hard. Their cell wall is covered by a layer of lignin which is water proof coating. Such seeds take more time for germination.
- The dormancy of seeds break by different artificial methods, like
- By making minute hole/pores with help of pointed sharp apparatus on the seed coat.
- The seeds are rubbed on hard object so that seed coat become thin.
- The partial degeneration of seed coat is carried by the action of sulphuric acid.
Under natural conditions, seed dormancy is gradually overcome by the action of microbes (bacteria) in soil in the alimentary canal of fruit eater birds and due to the presence of high cold or high temperature.
DORMANT EMBRYO
- In many species, although, the embryo is completely not matured, when the seed ripen, even then it fails to germinates whenever the environmental conditions are favourable or even seed coat is removed. This is known as "embryo dormancy".
- Embryo dormancy occurs in many fruits yielding plants of forest. This condition is achieved due to physiological action of seed. Such type of seed must complete their enzymatic and chemical reaction before the germination of seed. In the lack of these reactions, seeds are unable to germinate, such seed are kept in low temperature and in desirable moisture. Due to this, seed dormancy can be broken.
- The low temperature is the main reason for germination of seeds of apple, peaches, pears, maple and pine.
GERMINATION INHIBITORS
- Germination of some seeds is sometimes checked or prevented by the presence of some chemical compounds which are called "germination inhibitors" such as ferulic acid present in tomato juice, caumarin, abscisic acid, dormin and para ascorbic acid etc.
- This condition is generally found in xerophytic plants. These germination inhibitors washed away with water is known as "leaching".
FRUIT FORMATION
- Fertilized ovary with or without a seed together with any accessory structures closely associated with them is called fruit.
- Pomology is the study of fruits.
- On the basis of formation of fruits, they are classified into - true fruits and false fruits.
- The fruits which are developed only from the ovary and in which no other floral parts takes part in its development are called true fruits.
- The fruits in which addition to ovary other floral parts also contributes for its development are called false fruits. E.g., aggregate fruits.
- A fruit that is formed without fertilization & thus without seed is called seedless fruit or parthenocarpic fruits, e.g., banana, guava, apple, etc.
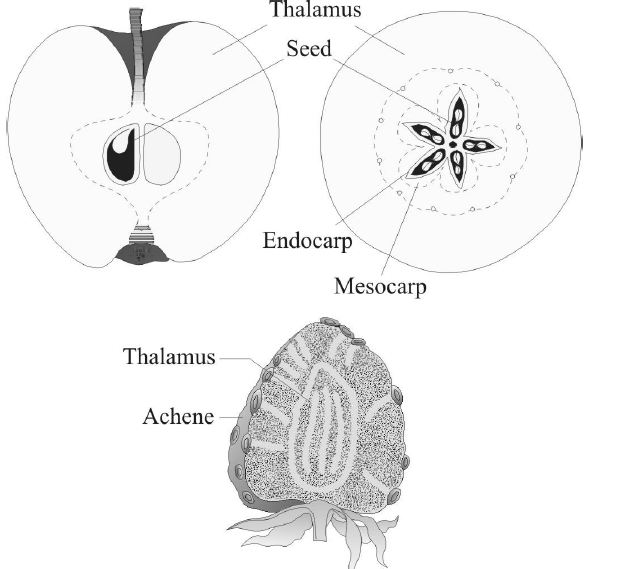
Fig. : False fruits of apple and strawberry
APOMIXIS, PARTHENOCARPY, POLYEMBRYONY
- Apomixis is a modified form of asexual reproduction in which seeds are formed without fusion of gametes. It includes the process whereby a diploid cell of the nucellus develops into an embryo, giving a diploid seed with a genetic constitution identical to the parent.
- Apomixis is usually associated with polyploidy. The organism that can reproduce by apomixis is called an apomict. E.g., dandelions, citris and certain grasses.
- Parthenocarpy is the development of fruit in an unfertilized flower, resulting in a seedless fruit. It may occur naturally as in certain varieties of pineapple, grapes, apple, pear and banana.
- Polyembryony is the presence of more than one embryo in the seed. It may be due to the presence of more than one egg cell in the embryo sac or more than one embryo sac in the ovule, and all the egg cells may get fertilized.
- Many embryos are formed inside the single seed is called Polyembryony. It was observed by Leeuwenhoek in Citrus (Orange) seeds.
- Polyembryony is commonly found in gymnosperms but it is also found in some of angiospermic plants such as orange, lemon and Nicotiana, etc.
- If polyembryony develops naturally then it is called spontaneous polyembryony (self), while if it develops artificially it is called induced polyembryony.
- When many embryos are formed from separate-separate embryo sacs (more than one) inside the ovule it is called false or pseudopolyembryony.
- When many embryos are formed inside the single embryo sac of the seed it is called "true polyembryony". It either develops from :
- Cleavage of zygote or budding : E.g., Cymbidium, Exocarpus, Nymphaea, Nicotiana, Orchids etc.,
- By the fertilization of synergids : E.g., Anemone, Aristolochia, Sagittaria, etc.
- Fertilization of antipodal cells : E.g., Paspalum, Ulmus, etc.
NOTES
- In Lilium, all three types of polyembryony are present.
- Adventive embryony is also an example of polyembryony in which additional number of embryos are formed from nucellus or integuments.
Study Notes for NEET/AIIMS/JIPMER




